 "
"
Team:Imperial College London/Drylab/Enzyme/Analysis/Detailed
From 2009.igem.org
(→Equation 4-MM: New rate of change of product) |
|||
| (3 intermediate revisions not shown) | |||
| Line 52: | Line 52: | ||
| - | <b>Equation 4-MM: New rate of change of product</b> | + | ====<b>Equation 4-MM: New rate of change of product</b>==== |
[[Image:ii09_ekeqnk4mm1.png | 100px]] | [[Image:ii09_ekeqnk4mm1.png | 100px]] | ||
| Line 70: | Line 70: | ||
[[Image:ii09_enzymelegend.png]] | [[Image:ii09_enzymelegend.png]] | ||
| + | ===References=== | ||
| + | [1]Steven H. Strogatz (1994). Nonlinear dynamics and chaos: with applications to physics, biology chemistry and engineering. Addison Wesley. ISBN 0-201-54344-3<br> | ||
| + | [2]Physica A: Statistical and Theoretical Physics, Vol. 188, No. 1-3. (1 September 1992), pp. 404-425. A rigorous derivation of the chemical master Equation | ||
{{Imperial/09/TemplateBottom}} | {{Imperial/09/TemplateBottom}} | ||
Latest revision as of 08:42, 14 October 2009

The behaviour of the system (the single-substrate mechanism) can be described with four ordinary differential equations (ODEs). Each component is described by a simple rate term for each of the four species involved in the mechanism. The rate term is a direct consequence of the application of the law of mass action.
We adopt the following notations for the kinetic rate constants involved in the single-substrate mechanism
The Enzyme Mechanism

In substrate binding, 1 molecule of Enzyme (E) binds with 1 molecule of Substrate (S) in a reversible reaction to form the Enzyme-Substrate complex (ES). From an experimental point of view, this step is blind: we are unable to measure this step. In catalysis, the Enzyme-Substrate complex (ES) produces the unchanged enzyme (E) and the product (P) in an irreversible reaction. This is the step that can be measured experimentally. When [S] becomes high enough, the second step of the enzyme kinetics becomes the rate determining step, controlled by k3. (link to simulation results)
Equation 1: Rate of change of Enzyme

Equation 2: Rate of change of substrate

Without MM assumption
Equation 3: Rate of change of enzyme-substrate complex

Equation 4: Rate of change of product

With Michaelis-Menten Assumptions
Consider the enzyme mechanism again:
From the model, we want to derive an equation that describes the rate of enzyme activity (amount of product formed per time interval) as a function of substrate concentration.
Consider equation 4: the rate of product formation. This equation is essentially useless as we cannot measure [ES] experimentally, therefore, to correctly determine d[P]/dt, we need to solve for ES in terms of the other quantities.
Therefore, we need to make some approximations as mentioned previously. (hyperlink to assumptions part)
At steady state, equation 3 is reduced to:
![]() When this result is used together with equation 4,
a new differential equation for rate of change of product is obtained:
When this result is used together with equation 4,
a new differential equation for rate of change of product is obtained:
Equation 4-MM: New rate of change of product
![]()
Michaelis-Menten assumption:
- The Michaelis-Menten (MM) assumption is a simplifying assumption that is commonly made with enzymatic reactions. It says that since substrate binding is very fast compared to catalysis, the enzyme complex ES is always at quasi-steady state, ie:
 , during the time of the experiment.
, during the time of the experiment.
- It is straightforward to prove that combining the MM assumption
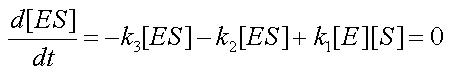 with the conservation equation [E0]=[E]+[ES] yields a new form for the fourth equation
with the conservation equation [E0]=[E]+[ES] yields a new form for the fourth equation 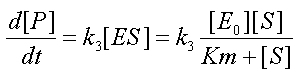 , where KM is the Michaelis-Menten constant.
, where KM is the Michaelis-Menten constant.


Equation (4-MM) seems more complex but is in practice more useful exploitable than Equation (4). Indeed, [S] can often be measured during an experiment, whereas [ES] cannot. Furthermore when the initial concentration of substrate [S0] is large compared to KM, then [S] remains large compared to KM over a certain range of time (that includes the start of the assay) and (4-MM) simplifies to ![]() over this range of time. Simply put, for these times P is produced at a constant rate at the start of the assay. Because of its close association to one of the possible behaviours of the system, the KM is one of the important pieces of information that can be given about an enzymatic reaction.
over this range of time. Simply put, for these times P is produced at a constant rate at the start of the assay. Because of its close association to one of the possible behaviours of the system, the KM is one of the important pieces of information that can be given about an enzymatic reaction.
Legend

References
[1]Steven H. Strogatz (1994). Nonlinear dynamics and chaos: with applications to physics, biology chemistry and engineering. Addison Wesley. ISBN 0-201-54344-3
[2]Physica A: Statistical and Theoretical Physics, Vol. 188, No. 1-3. (1 September 1992), pp. 404-425. A rigorous derivation of the chemical master Equation


