 "
"
 Alternative way of binding: Jun/Fos
Alternative way of binding: Jun/Fos
 |
| Figure1: Human c-Fos:c-Jun:DNA complexes |
Introduction
We also thought of an alternative way of binding of the heterodimeric Fok to the DNA avoiding the necessity of labeled oligos and their binding proteins. Therefore, we used the DNA binding domain of a transcription factor as a new adapter between the Fok construts and the DNA. We focused on the binding domain of the Activator Protein-1 (AP-1), a crucial transcription factor implicated in numerous cancers.
AP-1, which belongs to the bZIP type of transcription factors, binds DNA as a dimer like many eukaryotic transcription factor. Protein-protein interaction is mediated via a leucin zipper (ZIP), and with their basic region (b) they bind DNA. Nine homologues of the AP-1 leucine zipper region have been characterized, among them natural occurring c-Fos, c-Jun and library-designed winning peptides FosW and JunW. Each of the homologues is able to form heterodimers, some also form homodimers.
There are two classes of core DNA sequences, the sequences TRE (TGACTCA) and CRE (TGACGTCA), which are recognized by the different AP-1 factors.
For our needs, we chose the natural bZIP sequence of Fos and Jun as well as the library-selected FosW sequence. By fusion of Fos with Splitli-Fok_a, the construct Fos-Splitli-Fok_a was obtained. Control of DNA cleavage occurs on two levels: (i) using the bZIP sequence of Jun as adapter, DNA binding can be induced, and (ii) adding Fok_i will render Fok activ leading to DNA cleavage.
For further control of DNA cleavage activity, a light-switchable peptide, FosW, can be used to interfere with Fos/Jun DNA binding. For this, two Cys residues were introdued in FosW presenting reactive sites for thiol-reactive linkers. Thus an azobenzene derivative as intramolecular cross-linker can be coupled to the cysteines. Depending on the wavelenth of light used the linker undergoes cis/trans isomerization promoting the folding or unfolding of the FosW helix (see Figure 3).
 |
| Figure 3: light-switchable FosW interrupts binding of Jun:Fos |
Using this light-switchable FosW derivative, FosW binds in its light-activated form to Jun and Fos, displacing them from the DNA and thus disrupts Fok-mediated DNA cleavage. Switching FosW back to its inactivated form, Jun and Fos are released, can rebind to the DNA and Fok-mediated cleavage should occur again.
The assay with the photoswitchable FosWinner interrupting the binding of the heterodimeric complex of Fos and Jun was already done by the laboratory of Katja Arndt in Freiburg and underlines the feasibility of the method.
Methods
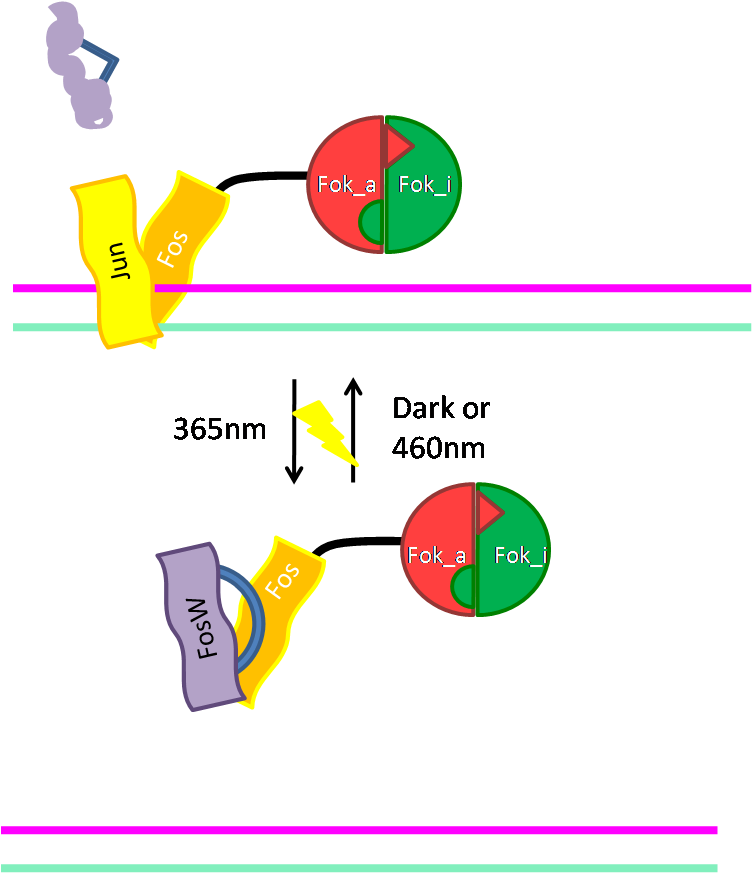 |
| Figure 2: Above: Fos and Jun dimerize, bind the DNA and lead Fok construct to DNA, Below: activated FosW prevents DNA binding |
Results and Discussion
The cloning of the construct His_Fos and Splitli-Foka into the pEX expression vector was done by a triple ligation. The first insert, His_Fos was cut out by XbaI and AgeI, the second insert, Splitli-Fok_a by NgoMIV and PstI, the vector was cut open with XbaI and PstI. The cloning was successful, and the concentration of the plasmid and the sequencing of clones were good.
| Sample ID | User ID | Date | Time | ng/ul | A260 | A280 | 260/280 | 260/230 | Constant | Cursor Pos. | Cursor abs. | 340 raw |
| pExFosSplitFokaK1 | FreiGEM | 14.10.2009 | 19:36 | 76.19 | 1.524 | 0.758 | 2.01 | 2.16 | 50.00 | 230 | 0.705 | -0.009 |
| pExFosSplitFokaK2 | FreiGEM | 14.10.2009 | 19:37 | 50.13 | 1.003 | 0.491 | 2.04 | 2.21 | 50.00 | 230 | 0.453 | 0.078 |
The recombinant expression of the single parts His_Fos-Splitli-Fok_a, GST-FosW and His-FluA-Splitli-Fok_i was performed. His-FluA-Splitli-Fok_i was successfully purified via a Ni-NTA column. The western blot with an anti-His antibody shows a band of the expected size of 467 kDa in the pooled elution fraction. GST-FosW was expressed and coupled to the GSH-sepharose column for the purification of His_Fos-Splitli-Fok_a, which will be the next step followed by the in vitro cleavage assays.
|
| Figure 5: Western Blot (anti-His antibody): His-Flu_a-Split_Fok_i in pEx; lanes: NEB prestained marker broad range, pool of elution fractions 2-5, empty lane, 3 positive controls for His-tag detection. |